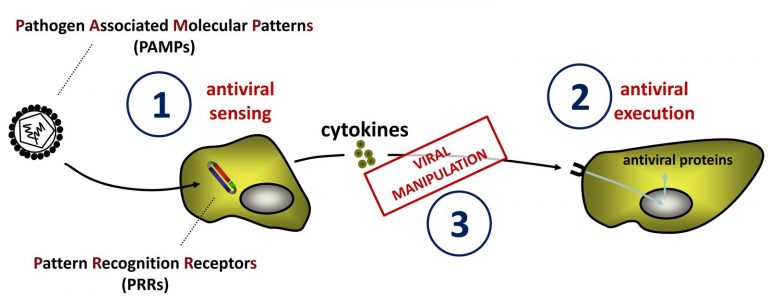
(1) Interaction of viral nucleic acids with host proteins
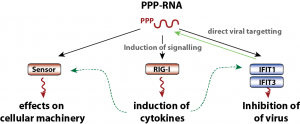
We identified viral triphosphorylated RNA as specific ligand for the virus sensor RIG-I (Pichlmair et al., Science 2006). Using affinity proteomics followed by mass spectrometry we identified additional proteins binding specifically to this type of RNA. Interferon induced proteins with tetratricopeptide repeats (IFIT), for instance, bind PPP-RNA and perturb virus growth (Pichlmair et al, Nature Immunology, 2011). IFIT proteins bind PPP-RNA using a uniqe mechanism ensuring high specificity and affinity (Abbas et al., Nature 2013). IFIT1 depletion in vitro and in mice are specifically susceptible to infection with viruses including orthomyxo- (e.g. influenza A virus) and paramyxoviruses (e.g. vesicular stomatitis virus). Functionally, IFITs specifically target translation of viral RNA (Habjan et al., Plos Pathogens, 2013).
Using similar approaches we identified a yet unstudied protein, NCBP3, as cap-binding protein (Gebhardt et al., Nature Communications 2015). NCBP3 binds NCBP1 to from an alternative Cap-RNA complex (CBC) that binds to mRNA and is important for RNA processing and export. Lack of NCBP3 is increasing vulnerability to virus infections, suggesting an important role of the alternative CBC during antiviral responses.